
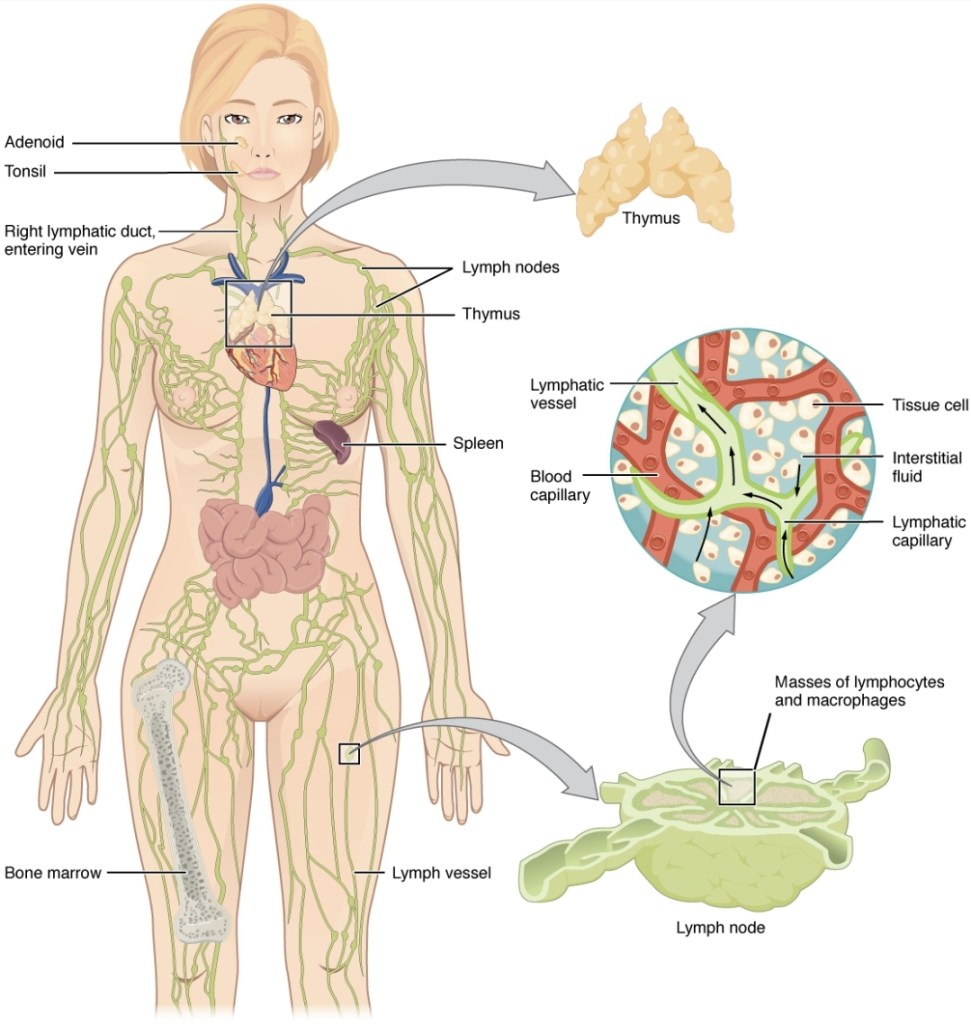
Nicień , Źródło: scienceblogs.com
Patolog Dr Alan MacDonald wykrył nicienie podczas autopsji tkanek mózgu pacjentów chorych na borelioze Lyme, stwardnienie rozsiane, demencje, glejaka mózgu oraz chorobę Alzheimera. Dr MacDonald wykazał również iż krętki borrelia ukrywają się we wnętrzu pasożytniczych nicieni, powodując przewlekłe choroby mózgu ,znalazł on trzy szczepy bakterii borrelia żyjących w pasożytniczych nicieniach, jajach nicieni oraz larwach tych pasożytów . MacDonald twierdzi, że zarówno nicienie jak i borrelia mogą powodować katastrofalne uszkodzenie mózgu.

Zdjęcia przedstawiają nicienia ,bakterie borrelia w jego wnętrzu oraz symbiotyczną bakterie Wolbachia.
Źródło: Prezentacja Dr Alan McDonald

Wektorem przenoszącym nicienie (Nematodes) może być kleszcz, komar oraz mucha.
Gdy już nicień zainfekuje gospodarza , uwalnia do układu krwionośnego tysiące larw zwanych mikrofilariami. Kiedy mikrofilarie dojrzewają do postaci larwy , uszkadzają mózg i powodują wiele objawów neurologicznych. Larwy nicieni mogą uszkodzić tkanki mózgu , powodować zwłóknienie, zatory w naczyniach krwionośnych mózgu, , miejscowe stany zapalne , osłabienie, zaburzenia widzenia. Nicienie manipulują układem immunologicznym w celu budowania przyjaznego dla siebie ekosystemu.U chorych zainfekowanych nicieniem diagnozuje się podwyższone poziomy markerów zapalnych takich jak czynnik wzrostu śródbłonka naczyniowego (VEGF), czynnik wzrostu hepatocytów (HGF) oraz metaloproteinazy macierzy 9 (MMP-9). Infekcja nicieniem może być mylona z objawami bartonella. Eliminowanie nicieni farmaceutykami oraz ziołami może prowadzić do silnej reakcji Herxheimera. U chorych na stwardnienie rozsiane reakcja herxaimera może być szczególnie groźna. Źródłem reakcji herxaimera nie jest jednak sam nicien lecz Wolbachia , symbiotyczna bakteria żyjąca w nicieniu. Wolbachia należy do Alfaproteobakterii podobnie jak Bartonella, Brucella , Rickettsia oraz Anaplasma. Gdy nicienie są zabijane przez leki lub zioła przeciwpasożytnicze, Wolbachia nie mogąc przeżyć bez swojego żywiciela ginie, uwalniając swoje endotoksyny. Endotoksyny Wolbachia stymulują cytokiny prozapalne IL-6, TNF-alfa,IL-1beta,IL-12 powodując silny stan zapalny. Dodatkowo Wolbachia żyjąc w nicieniu inicjuje procesy prozoaplne , aktywuje cytokine IL-8( jeden z najsilniej działających na neutrofile czynników chemotaktycznych) oraz hamuje apoptozę neutrofili. Wysoki poziom neutrofili może wskazywać na jej obecność. Należy podkreślić iż Wolbachia steruje dodatkowo procesami życiowymi nicieni , neutralizowanie białka powierzchniowego tej bakterii może zakłócić cykl replikacyjny oraz być jedną ze strategii eliminowania nicieni. Kluczowe również jest hamowanie samej cytokiny IL-8 która inicjuje stany zapalne.

Wolbachia , Źródło: medicineworld.org
Reasumując nicienie same w sobie są w stanie powodować uszkodzenia mózgu , symbiotyczna bakteria Wolbachia inicjuje dodatkowe stany zapalne oraz reakcje herx ,nicień jest również swego rodzaju biofilmem dla krętka borrelia. Aby skutecznie eliminować nicienie z układu nerwowego oraz mózgu konieczna jest kombinacja ziół, które wyeliminują zarówno larwy jak i mikrofilarie , zneutralizują bakterie Wolbachia oraz zredukują poziom cytokiny prozapalnej IL-8. Eliminowanie niecni pokrywa się w pewnym stopniu z neutralizowaniem bartonella.
Zioła i olejki eteryczne (oleum aetherium) eliminujące mikrofilarie
Andrographis paniculata,Olejek goździkowy (Syzygium aromaticum)
Zioła i olejki eteryczne eliminujące larwy
Olejek goździkowy(Syzygium aromaticum), Olejek tymiankowy(Thymus vulgaris), Olejek palmarozowy(Cymbopogon martini), Lantana camara, Ocimum sanctum

Lantana pospolita (Lantana camara L.)
Zioła i olejki eteryczne neutralizujące białka powierzchniowe wolbachia
Olejek kajeputowy z kwiatów Melaleuca cajuputi – znacznie zmniejsza endosymbiont Wolbachia w komórkach Aa23, białka powierzchniowego Wolbachia, co prowadzi do hamowania uwolnienia mikrofilarii, a także skrócenia żywotności i ruchliwości dorosłych nicieni.

Melaleuca cajuputi
Zioła i olejki eteryczne neutralizujące cytokine IL-8
Ailanthus (Bai Chun PI), Capillaris (YinChen Hao), Cordyceps (Dong Chong Xia Cao), Coriolus (Yun Zhi), curcuma longa (Jiang Huang), Ganoderma (Ling Zhi), Magnolia Kora (Hou Po), Silybum (Chai Hu), Mahonia (Lao Gong Ye)Moutan (Dan Pi), Szałwia czerwona (Dan Shen), Vitex (Man Jing Zi), Olejek palmarozowy(Cymbopogon martini)

Mahonia
Zioła i olejki eteryczne redukujące cytokiny prozapalne IL-6, TNF-alfa,IL-1beta,IL-12, VEGF, MMP-2, MMP-9
Olejek goździkowy(Syzygium aromaticum) redukuje VEGF, MMP-2, MMP-9
Olejek palmarozowy(Cymbopogon martini) redukuje TNF-alfa, IL-1ß, IL-8 oraz aktywuje IL-10.
Olejek tymiankowy(Thymus vulgaris) redukuje IL-1beta, IL-6, GM-CSF, TNFalpha
Andrographis redukuje IL-1, IL-1ß, IL-6 , MMP-2, IL-1beta , VEGF.
Zioła i suplementy redukujące reakcje Herxheimera :
Melatonina, Glutation , Tarczyca bajkalska(Scutelleria)

Scutelleria
Nie stosujemy olejków eliminujących nicienie bez ziół redukujących reakcje Herx.
Najkorzystniej jest zacząć od ziół neutralizujących białko powierzchniowe Wolbachia oraz hamujących cytokiny prozapalne.
U chorych na Stwardnienie rozsiane zaczynamy od bardzo niewielkich dawek ziół stopniowo je zwiększając. W momencie wystąpienia reakcji Herxheimera zmniejszamy dawki ziół lub odstawiamy zioła na 3-4 dni .
UWAGA: Informacje na tej stronie służą wyłącznie celom edukacyjnym i nie mogą być używane w celach diagnostycznych lub terapeutycznych. Informacje te nie mogą być stosowane jako substytut profesjonalnego diagnozowania i leczenia. Przed podjęciem jakiejkolwiek terapii, należy skonsultować się z lekarzem lub fitoterapeutą .
______________________________________________________________________________________
Bibliografia:
M.W. Kennedy, W. Harnett (Eds.): Parasitic Ne
matodes: Molecular Biology, Biochemistry and
Immunology. CABI Publishing, Oxon and New York,
2001. ISBN 0-85199-423-7,

Korean J Parasitol. 2016 Jun;54(3):273-80. doi: 10.3347/kjp.2016.54.3.273. Epub 2016 Jun 30.
Antifilarial and Antibiotic Activities of Methanolic Extracts of Melaleuca cajuputi Flowers.
Al-Abd NM, Nor ZM, Mansor M, Hasan MS, Kassim M.
Parasite Immunol. 2007 Feb;29(2):73-9.
Wolbachia surface protein (WSP) inhibits apoptosis in human neutrophils.
Bazzocchi C, Comazzi S, Santoni R, Bandi C, Genchi C, Mortarino M.
Immunological role of the endosymbionts of Dirofilaria immitis: the Wolbachia surface protein activates canine neutrophils with production of IL-8.
Bazzocchi C, Genchi C, Paltrinieri S, Lecchi C, Mortarino M, Bandi C.
Vet Parasitol. 2003 Nov 3;117(1-2):73-83.
Kamaraj, Chinnaperumal, Abdul Abdul Rahuman, Gandhi Elango, Asokan Bagavan, and Abdul Abduz Zahir. “Anthelmintic Activity of Botanical Extracts against Sheep Gastrointestinal Nematodes, Haemonchus Contortus.” Parasitology Research 109, no. 1 (July 2011): 37–45. doi:10.1007/s00436-010-2218-y.
Merawin, L. T., A. K. Arifah, R. A. Sani, M. N. Somchit, A. Zuraini, S. Ganabadi, and Z. A. Zakaria. “Screening of Microfilaricidal Effects of Plant Extracts against Dirofilaria Immitis.” Research in Veterinary Science 88, no. 1 (February 2010): 142–47. doi:10.1016/j.rvsc.2009.05.017.
Zaridah, M. Z., S. Z. Idid, A. W. Omar, and S. Khozirah. “In Vitro Antifilarial Effects of Three Plant Species against Adult Worms of Subperiodic Brugia Malayi.” Journal of Ethnopharmacology 78, no. 1 (November 2001): 79–84.
Dutta, A., and N. C. Sukul. “Filaricidal Properties of a Wild Herb, Andrographis Paniculata.” Journal of Helminthology 56, no. 2 (June 1982): 81–84.
Raj, R. K. “Screening of Indigenous Plants for Anthelmintic Action against Human Ascaris Lumbricoides: Part–II.” Indian Journal of Physiology and Pharmacology 19, no. 1 (March 1975): UNKNOWN.
Salim, Emil, Endang Kumolosasi, and Ibrahim Jantan. “Inhibitory Effect of Selected Medicinal Plants on the Release of pro-Inflammatory Cytokines in Lipopolysaccharide-Stimulated Human Peripheral Blood Mononuclear Cells.” Journal of Natural Medicines 68, no. 3 (July 2014): 647–53. doi:10.1007/s11418-014-0841-0.
Tangyuenyong, Siriwan, Nawarat Viriyakhasem, Siriporn Peansukmanee, Prachya Kongtawelert, Siriwan Ongchai, Siriwan Tangyuenyong, Nawarat Viriyakhasem, Siriporn Peansukmanee, Prachya Kongtawelert, and Siriwan Ongchai. “Andrographolide Exerts Chondroprotective Activity in Equine Cartilage Explant and Suppresses Interleukin-1β-Induced MMP-2 Expression in Equine Chondrocyte Culture, Andrographolide Exerts Chondroprotective Activity in Equine Cartilage Explant and Suppresses Interleukin-1β-Induced MMP-2 Expression in Equine Chondrocyte Culture.” International Scholarly Research Notices, International Scholarly Research Notices 2014, 2014 (October 30, 2014): e464136. doi:10.1155/2014/464136, 10.1155/2014/464136.
Zhao, Feng, En-Qi He, Lu Wang, and Ke Liu. “Anti-Tumor Activities of Andrographolide, a Diterpene from Andrographis Paniculata, by Inducing Apoptosis and Inhibiting VEGF Level.” Journal of Asian Natural Products Research 10, no. 5–6 (June 2008): 467–73. doi:10.1080/10286020801948334.
Begum, Sabira, Syeda Qamar Zehra, Bina Shaheen Siddiqui, Shahina Fayyaz, and Musarrat Ramzan. “Pentacyclic Triterpenoids from the Aerial Parts of Lantana Camara and Their Nematicidal Activity.”Chemistry & Biodiversity 5, no. 9 (September 2008): 1856–66. doi:10.1002/cbdv.200890173.
Misra, Namita, Mithilesh Sharma, Kanwal Raj, Anil Dangi, Sudhir Srivastava, and Shailja Misra-Bhattacharya. “Chemical Constituents and Antifilarial Activity of Lantana Camara against Human Lymphatic Filariid Brugia Malayi and Rodent Filariid Acanthocheilonema Viteae Maintained in Rodent Hosts.” Parasitology Research 100, no. 3 (February 2007): 439–48. doi:10.1007/s00436-006-0312-y.
Santana, Omar, Maria Fe Andrés, Jesús Sanz, Naima Errahmani, Lamiri Abdeslam, and Azucena González-Coloma. “Valorization of Essential Oils from Moroccan Aromatic Plants.” Natural Product Communications 9, no. 8 (August 2014): 1109–14.
Giarratana, F., D. Muscolino, C. Beninati, A. Giuffrida, and A. Panebianco. “Activity of Thymus Vulgaris Essential Oil against Anisakis Larvae.” Experimental Parasitology 142 (July 2014): 7–10. doi:10.1016/j.exppara.2014.03.028.
Kumaran, Asha M., Prashanth D’Souza, Amit Agarwal, Rama Mohan Bokkolla, and Murali Balasubramaniam. “Geraniol, the Putative Anthelmintic Principle of Cymbopogon Martinii.” Phytotherapy Research: PTR 17, no. 8 (September 2003): 957. doi:10.1002/ptr.1267.
Katiki, L. M., A. C. S. Chagas, H. R. Bizzo, J. F. S. Ferreira, and A. F. T. Amarante. “Anthelmintic Activity of Cymbopogon Martinii, Cymbopogon Schoenanthus and Mentha Piperita Essential Oils Evaluated in Four Different in Vitro Tests.” Veterinary Parasitology 183, no. 1–2 (December 29, 2011): 103–8. doi:10.1016/j.vetpar.2011.07.001.
Duarte, Marta Cristina Teixeira, Ewerton Eduardo Leme, Camila Delarmelina, Andressa Almeida Soares, Glyn Mara Figueira, and Adilson Sartoratto. “Activity of Essential Oils from Brazilian Medicinal Plants on Escherichia Coli.” Journal of Ethnopharmacology 111, no. 2 (May 4, 2007): 197–201. doi:10.1016/j.jep.2006.11.034.
Khan, Mohd Sajjad Ahmad, and Iqbal Ahmad. “In Vitro Antifungal, Anti-Elastase and Anti-Keratinase Activity of Essential Oils of Cinnamomum-, Syzygium- and Cymbopogon-Species against Aspergillus Fumigatus and Trichophyton Rubrum.” Phytomedicine: International Journal of Phytotherapy and Phytopharmacology 19, no. 1 (December 15, 2011): 48–55. doi:10.1016/j.phymed.2011.07.005.
Tsai, Mei-Lin, Chih-Chien Lin, Wei-Chao Lin, and Chao-Hsun Yang. “Antimicrobial, Antioxidant, and Anti-Inflammatory Activities of Essential Oils from Five Selected Herbs.” Bioscience, Biotechnology, and Biochemistry 75, no. 10 (2011): 1977–83.
Murbach Teles Andrade, Bruna Fernanda, Bruno José Conti, Karina Basso Santiago, Ary Fernandes Júnior, and José Maurício Sforcin. “Cymbopogon Martinii Essential Oil and Geraniol at Noncytotoxic Concentrations Exerted Immunomodulatory/anti-Inflammatory Effects in Human Monocytes.” The Journal of Pharmacy and Pharmacology 66, no. 10 (October 2014): 1491–96. doi:10.1111/jphp.12278.
Meyer, Susan LF, Dilip K Lakshman, Inga A Zasada, Bryan T Vinyard, and David J Chitwood. “Dose–response Effects of Clove Oil fromSyzygium Aromaticum on the Root-Knot nematodeMeloidogyne Incognita.” Pest Management Science 64, no. 3 (March 2008): 223–29. doi:10.1002/ps.1502.
Meyer, Susan LF, Dilip K Lakshman, Inga A Zasada, Bryan T Vinyard, and David J Chitwood. “Dose–response Effects of Clove Oil from Syzygium Aromaticum on the Root-Knot Nematode Meloidogyne Incognita.” Pest Management Science 64, no. 3 (March 2008): 223–29. doi:10.1002/ps.1502.
Manikandan, Palrasu, Ramalingam Senthil Murugan, Ramamurthi Vidya Priyadarsini, Govindarajah Vinothini, and Siddavaram Nagini. “Eugenol Induces Apoptosis and Inhibits Invasion and Angiogenesis in a Rat Model of Gastric Carcinogenesis Induced by MNNG.” Life Sciences 86, no. 25–26 (June 19, 2010): 936–41. doi:10.1016/j.lfs.2010.04.010.













 Źródło :
Źródło : 

